人类祖先留下的这面“免疫盾牌”
如果有人告诉你:一种如今被认为能够帮助人类抵抗HIV的关键基因突变,可能早在约9000年前就已经出现;而它最初的产生,并不是为了对抗HIV,因为HIV与人类的接触大约始于20世纪早期,直到1981年人类才首次正式识别并报告艾滋病。
这样看起来,这种古老的基因变异仿佛并非为现代病毒而生,却在数千年后意外成为人类对抗HIV的一道天然屏障。听起来,是否带着一种跨越时空的奇妙巧合?

但这正是一项发表于国际顶级期刊《细胞》(Cell)的最新研究给出的答案。
研究人员通过现代基因组学和古DNA技术,追踪到一种著名基因突变CCR5Δ32的起源,发现它最早可能出现在约6700—9000年前的黑海附近地区。
更重要的是,今天许多携带这一突变的人,可能都能把自己的遗传线索,追溯到那位远古祖先。
这项研究不仅讲述了一个“跨越千年的基因故事”,也在提醒我们:人类今天面对疾病时所依赖的一些“天然防线”,也许并不是为现代疾病而准备的,而是远古演化留下的意外馈赠。
神奇盾牌是什么?
那么,这个基因究竟是什么?它在人体中发挥着怎样的作用?又为什么某种突变,竟然能成为抵御病毒的一道“天然屏障”?
经历过新冠疫情之后,很多人对病毒感染人体的过程,已经有了一个相对直观的认识:病毒并不是“凭空钻进”细胞里的,它往往需要先识别并结合宿主细胞表面的特定分子,才能打开入侵的大门。
当然,不同病毒进入细胞的方式并不完全相同,但对许多病毒而言,感染的第一步,通常都离不开与宿主细胞表面的某种受体、辅助受体,或者附着分子发生结合。只有完成这一步,病毒才可能进一步进入细胞,并启动后续复制过程。CCR5(C-C chemokine receptor type 5),就是这样一个关键分子。
从生物学上说,CCR5是一种存在于部分免疫细胞表面的趋化因子受体,主要分布在某些T细胞、巨噬细胞等细胞上,参与免疫细胞迁移、炎症反应调控等过程。
也就是说,它本来就是人体免疫系统正常运转的一部分,并不是专门为病毒“准备”的。
但问题在于,病毒往往会“借用”人体原本就存在的通路。对于许多HIV-1毒株来说,CCR5恰恰就是它们进入宿主细胞时所依赖的重要辅助受体之一。
通常情况下,HIV-1会先与宿主细胞表面的CD4分子结合,随后再进一步结合CCR5,并由此促成病毒包膜与细胞膜融合,最终完成入侵。正因为如此,CCR5可以被理解为许多HIV-1毒株进入细胞时所依赖的一道关键“入口”。
这样一来,一个非常自然的问题就出现了:如果这道“入口”发生了变化,病毒是不是就没那么容易进来了?
答案是肯定的。现有的分子机制研究和临床观察都表明,对于那些依赖CCR5进入细胞的HIV-1毒株而言,只要CCR5的结构或功能发生改变,病毒的入侵效率就会明显下降。
其中最著名的一种变化,就是开头所提到的CCR5Δ32。这是一种发生在CCR5基因上的32个碱基对缺失突变。
这段缺失会导致蛋白编码发生移码,使生成的CCR5蛋白结构异常,难以正常加工并稳定表达在细胞表面。结果就是,细胞表面可供病毒利用的功能性CCR5显著减少。
对于那些原本依赖CCR5完成入侵的HIV毒株来说,这就意味着它们更难顺利结合宿主细胞,也更难完成后续膜融合与感染过程。
也正因此,CCR5Δ32才会受到如此广泛的关注。目前的研究认为,携带两个CCR5Δ32突变拷贝的个体,对多数依赖CCR5的HIV-1毒株具有较强的天然抵抗力;而只携带一个突变拷的个体,虽然通常并不会完全避免感染,但在一些情况下,可能表现出较低的易感性,或者更缓慢的疾病进展。
换句话说,CCR5Δ32并不是让人获得一种“绝对免疫”,更不是对所有HIV毒株都同样有效;但它确实通过改变病毒入侵所依赖的关键分子通路,为人体增加了一道天然防线。
也正是因为这一点,CCR5Δ32才不只是一个普通的基因变异。它让我们第一次如此清楚地看到:一个看似微小的遗传改变,竟然可以在分子层面重塑病毒与宿主之间的博弈关系,甚至影响一个人面对感染时的命运。
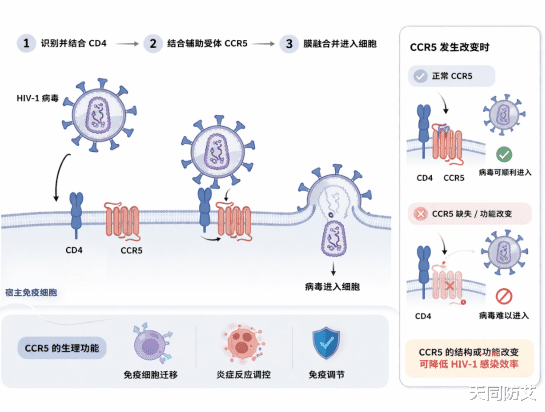
图1:HIV-1通过CD4和CCR5进入宿主细胞的示意图。
突变溯源:这段基因故事从哪里开始?
建立在大规模古今基因组分析基础上。研究团队共分析了934例古代基因组2504例现代基因组,结合古DNA、单倍型分析和概率模型,重建了CCR5Δ32的传播轨迹。结果表明,今天携带这一突变的人群,很可能都可以追溯到同一个古老的起源事件。换句话说,研究者像是在进行一场跨越数千年的“基因寻亲”:先在现代人中锁定这一特殊遗传标记,再到古代人类遗骸中寻找它最早出现的证据,最后将这条时间线一步步向更早时期推回。配合图中的时空分布可以更直观地看到,携带CCR5Δ32缺失的古代样本并非随机零散出现,而是在随时间推进的过程中,逐渐从较早期、较局部的分布扩展到更广泛的区域。这一证据链最终将该突变的源头指向了黑海附近、与西欧亚草原相关的新石器时代人群,说明这种今天因与抗HIV相关而备受关注的基因变异,其实早在数千年前就已经出现在人类祖先之中。
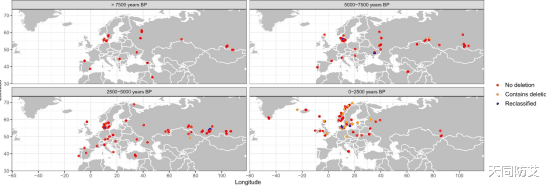
图2:CCR5Δ32基因分型的古代基因组地理分布
为什么这一突变能够被保留下来?
如果CCR5Δ32并不是为了对抗HIV才出现的,那么,它为什么会在人群中被保留下来,甚至在相当长一段时期内持续扩散?研究团队进一步对CCR5Δ32在西欧亚地区的时空频率变化进行了建模分析。
结果显示,这一突变的频率在距今约8000年至2000年之间经历了明显上升,随后才逐渐趋于稳定。更重要的是,统计模型提示,这种变化并不能仅用随机遗传漂变来解释,而更符合正向选择作用的结果。换句话说,在古代环境中,携带这一突变的人群,很可能确实获得了某种生存优势。
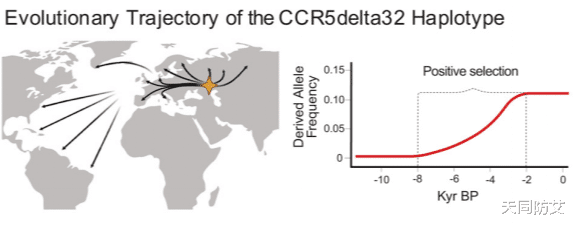
图3:CCR5Δ32的起源、扩散与正向选择
那么,这种优势究竟来自哪里?研究者提出了一个颇具说服力的解释:CCR5Δ32可能与免疫反应的调节有关。直觉上,人们往往会认为免疫系统越强越好,但事实并非如此。
免疫能力不足,固然难以抵御病原体;可如果免疫反应过于强烈,同样会对机体自身造成损伤,严重时甚至可能引发致命的炎症反应。也就是说,决定生存结局的,未必只是“免疫够不够强”,还包括“免疫是否足够平衡”。
从人类历史进程来看,这一解释也具有相当合理的背景。大约在距今数千年前,人类社会正经历从狩猎采集向农业定居生活的深刻转变。
随着聚落形成、人口密度增加以及人与动物接触方式改变,传染病传播的机会显著上升,新的病原压力也不断出现。在这样的环境下,像CCR5Δ32这样能够改变免疫应答方式的突变,或许并不是让个体完全“不生病”,而是让他们在感染发生时,不至于因免疫系统反应过度而承受更大代价,从而提高生存和繁衍的机会。
这也提示我们,自然选择保留下来的,未必是最“强”的免疫系统,而往往是更“合适”的免疫系统。 从这个角度看,CCR5Δ32的扩散并不是一个简单的“抗病基因流行”故事,而更像是人类在长期病原压力下,对免疫平衡不断调整后留下的一段演化印记。
讨论与反思
说到这里,必须提醒一句:CCR5Δ32并不是什么“完美突变”。虽然它确实可能带来某些保护作用,尤其因与HIV抵抗相关而备受关注;
但与此同时,它也绝非没有代价。现代研究表明,CCR5参与的并不只是HIV入侵过程,还与炎症调控、免疫细胞迁移以及免疫监视等多种生理过程密切相关。也就是说,当CCR5的功能发生改变时,受到影响的往往并不只是某一种疾病,而可能是一整套彼此关联的免疫反应网络。
正因如此,CCR5Δ32的影响始终具有明显的“双刃剑”特征。一方面,它可能在某些情况下带来保护效应;
但另一方面,也有研究提示,它可能与某些病毒感染风险升高有关,例如西尼罗河病毒感染风险增加。此外,它与部分免疫相关疾病之间的关系,也仍在持续研究和讨论之中。
这也正是为什么,2018年引发全球巨大争议的“基因编辑婴儿”事件,会让科学界如此警惕。除了知情同意、技术不成熟和脱靶效应等问题之外,更核心的担忧之一在于:即便某个基因改变看似能够带来某一方面的益处,也不能忽视它在整个生理系统中的复杂作用。
一个基因,从来都不是一个可以随意拨动的简单开关。你“关闭”的,可能不只是某一种风险,也可能是某种我们尚未完全理解、却一直维持着生命平衡的重要机制。
如果借用一个更直观的比喻,CCR5Δ32就像《三国演义》中藤甲兵所穿的藤甲:它刀箭难伤,看似是极强的防护;但穿上藤甲的同时,也意味着更惧怕火攻。基因变异也是如此——它带来的保护,往往并不是无条件、无代价的。
而这项研究的另一项重要意义,在于它还推翻了一个流传已久的旧假说。
过去,学界曾长期猜测,CCR5Δ32的扩散可能与中世纪欧洲黑死病或天花等大规模瘟疫有关。 这种解释之所以流行,是因为它听上去十分合理:当大瘟疫来袭,能够提高生存率的遗传变异,的确更有可能在后代中被保留下来。
然而,新研究提供的证据并不支持这一说法。研究显示,CCR5Δ32在人群中的频率变化到约2000年前已大体趋于稳定,这意味着它的主要扩散过程远早于中世纪瘟疫时期。
换言之,推动这一突变传播的关键选择压力,更可能来自更早的人群迁徙、生活方式转变以及疾病生态环境的变化,而不是黑死病本身。
这正是古DNA研究最有力量的地方:它不再只是让我们停留在“合理猜测”的层面,而是让我们第一次有机会,真正把一个基因变异重新放回历史现场去理解。
从黑海附近的新石器时代人群,到今天的HIV医学研究,CCR5Δ32的故事横跨了将近一万年。它提醒我们,基因并不是静止不动的一串代码,而是一部仍在延续的生命史。我们今天身体里的某些遗传特征,可能源自古代人群在特定环境压力下被自然选择保留下来的变异;而这些古老的演化结果,也许要到几千年后,才会在另一种疾病面前显现出新的意义。
这正是人类演化最令人震撼之处:自然选择从不“预判未来”,但它留下的痕迹,却可能在未来的某一天,突然展现出意想不到的价值。
因此,这项研究的意义,不只是解答了CCR5Δ32从何而来,更提醒我们:在基因编辑与精准医学迅速发展的今天,人类对基因的理解,仍远没有到可以“随意改写”的程度。一个看似有益的突变,背后可能隐藏着复杂的代价;一个今天能帮助我们抵御某种疾病的遗传变化,当年也许只是自然选择在另一种环境压力下保留下来的结果。
所以,面对基因,我们既需要科学探索的勇气,也需要对生命复杂性的敬畏。因为真正重要的,从来不是“找到一个万能基因”,而是逐渐理解:人类身体里的每一个变异,都是时间、环境、疾病与生存共同写下的答案。
参考文献:
Ravn K, et al. Tracing the evolutionary history of the CCR5delta32 deletion via ancient and modern genomes. Cell, 2025.
Stephens JC, et al. Dating the origin of the CCR5-Delta32 AIDS-resistance allele by the coalescence of haplotypes. Am J Hum Genet, 1998.
Alkhatib G, et al. CC CKR5: a RANTES, MIP-1alpha, MIP-1beta receptor as a fusion cofactor for macrophage-tropic HIV-1. Science, 1996.
Choe H, et al. The beta-chemokine receptors CCR3 and CCR5 facilitate infection by primary HIV-1 isolates. Cell, 1996.